


El virus de la rabia: un germen que se transporta en contravía

Autores
Orlando Torres Fernández Biólogo, Magister en Morfología, Máster en Psicobiología y Neurociencia Cognitiva, Doctor en Ciencias Biomédicas, Grupo de Morfología Celular, Subdirección de Investigación Científica y Tecnológica, Instituto Nacional de Salud.
Gerardo Santamaría Romero Biólogo, Máster en Neurociencias y Biología del Comportamiento, Grupo de Morfología Celular, Subdirección de Investigación Científica y Tecnológica, Instituto Nacional de Salud
Jeison Alexander Monroy Gómez Biólogo, Especialista en Proyectos de Investigación Científica y Tecnológica, Candidato a Magister en Neurociencias, Grupo de Morfología Celular, Subdirección de Investigación Científica y Tecnológica, Instituto Nacional de Salud Docente, Departamento de Ciencias Básicas, Institución Universitaria Escuela Colombiana de Rehabilitación.
Resumen
El virus de la rabia es un germen patógeno que durante miles de años se ha mantenido circulando en la naturaleza y produce una de las enfermedades que más aterroriza a los seres humanos debido a sus efectos dramáticos y su carácter letal. Su afinidad selectiva por el sistema nervioso hace que este agente infeccioso tenga características especiales muy diferentes a la de otros patógenos que regularmente afectan a la humanidad. Una de ellas es su mecanismo de ingreso y diseminación dentro del organismo.
Introducción:
Generalmente el virus de la rabia es inoculado mediante mordedura a través de la saliva de un animal infectado. Luego se introduce en las terminaciones de los nervios periféricos que llegan a la piel o los músculos estriados de las extremidades superiores o inferiores por ser las áreas más expuestas, por lo menos en los humanos, y a través de ellos es transportado inicialmente a la médula espinal y luego al encéfalo. A diferencia de muchos otros patógenos el virus de la rabia no viaja por el torrente sanguíneo (no se presenta viremia); tampoco ingresa por el tracto digestivo como sí ocurre por ejemplo con el virus de la hepatitis A o el poliovirus. El anatomista italiano Giovanni Morganini (1682-1771), considerado el padre de la anatomía patológica moderna, fue uno de los primeros en especular que la rabia sería producida por un germen transportado a través de los nervios y no a través de los vasos sanguíneos. Esto debido a las alteraciones en la sensibilidad que manifiestan algunas personas en el sitio de la mordida (Jackson, 2013:1-15).
En 1881 Luis Pasteur, pionero de la investigación de la rabia en tiempos modernos, junto con su equipo de trabajo, demostraron transmisión de la infección mediante inoculación intracerebral de conejos con extractos de cerebros de perros infectados. DiVestea y Zagari en 1889 inocularon el virus en el nervio ciático de perros y conejos. Los animales se enfermaron de rabia, no obstante, la enfermedad no se desarrolló cuando el nervio fue cortado y cauterizado. Goodpasture en 1925 logró, experimentalmente, enfermar animales mediante inoculación intramuscular del virus. Lo consiguió inyectando el músculo masetero de conejos. Sus observaciones lo llevaron a concluir que el virus pasó de las fibras musculares a los axones de uno de los nervios craneales y a través de ellos alcanzó el tallo cerebral (Jackson, 2013:1-15). A mediados del siglo XX, cuando ya el virus había sido aislado e identificado, se retomaron los experimentos en los cuales se inoculaba el virus en las extremidades posteriores de ratas, ratones y otros pequeños roedores, a los que previamente se les había cortado los nervios ciático y safeno. Se comprobó que este procedimiento evitaba el desarrollo de la enfermedad en la mayoría de estos animales. Además, siempre que se inoculaba el virus por vía intramuscular en las extremidades posteriores su presencia era detectada primero en la médula espinal antes que en el encéfalo (Schindler, 1961:119-126; Dean y otros, 1963:803-811).
Composición del sistema nervioso.
Para entender la ruta de diseminación del virus de la rabia es necesario hacer una breve descripción de las estructuras del sistema nervioso. Existen el sistema nervioso central (SNC) y el sistema nervioso periférico (SNP). El SNC corresponde al encéfalo y la médula espinal, es decir, las partes protegidas por el cráneo y las vértebras (Figura 1). El SNP lo componen principalmente nervios conectados con la médula espinal, pero localizados por fuera de las vértebras y distribuidos por todo el cuerpo. Los nervios periféricos motores tienen su origen en neuronas de gran tamaño (motoneuronas) ubicadas dentro del asta ventral de la médula espinal. Los axones de estas neuronas emergen ventralmente formando paquetes de nervios, algunos de ellos de gran calibre, como los que se dirigen hacia las extremidades posteriores (Figura 2).
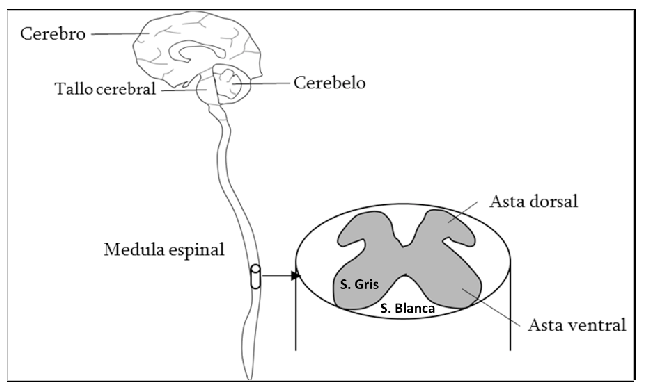
Figura 1. A la izquierda esquema de los componentes del Sistema Nervioso Central (SNC). A la derecha detalle de la distribución de la sustancia gris y la sustancia blanca dentro de la médula espinal.
Los nervios periféricos sensoriales son formados por axones de las neuronas de los ganglios espinales. Estos son pequeños abultamientos que se observan entre las uniones de las vértebras muy cerca de la médula espinal. De cada neurona de los ganglios espinales emerge un delgado proceso celular que se ramifica muy cerca y da lugar a dos ramas. Una de ellas es muy larga y se dirige hacia la periferia para recoger información sensorial de la piel y otros órganos que llega a su neurona de origen dentro del ganglio espinal, y luego la otra rama, mucho más corta conduce la información sensorial hasta el asta dorsal de la médula espinal (Figura 2). En resumen, los nervios periféricos motores llevan información desde la médula espinal hacia los músculos mientras que los nervios periféricos sensoriales conducen información sensorial desde la periferia hasta la médula espinal.
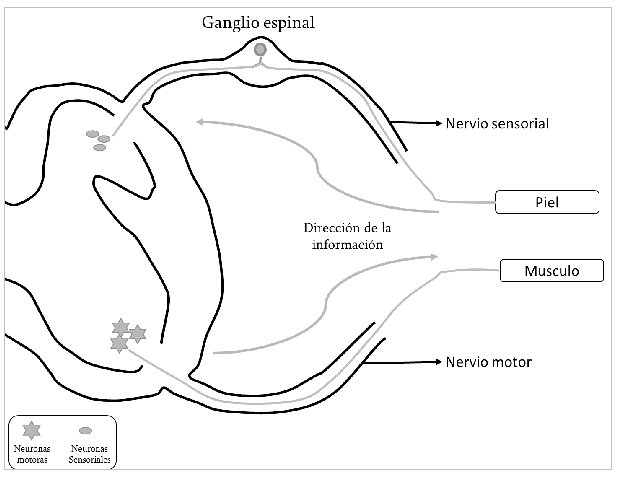
Figura 2. Origen de los nervios periféricos. Dentro de los ganglios espinales se encuentran las neuronas sensoriales de las cuales se desprenden dos ramas nerviosas: una recibe información desde la periferia y la otra la conduce hacia el asta dorsal de la médula espinal. En el asta ventral se encuentran las neuronas motoras; sus axones se dirigen a los músculos para provocar su contracción.
La médula espinal está conformada por sustancia gris (donde se encuentran los cuerpos de las neuronas) localizada hacia el centro en forma de mariposa y por sustancia blanca constituida por los tractos o fascículos que la rodean (Figura 1). Algunos de ellos están conformados por axones de neuronas con función motora (tractos descendentes) y otros por axones de neuronas con función sensorial (tractos ascendentes). Como regla general la información sensorial fluye desde los diferentes niveles medulares (sacro, lumbar, torácico y cervical) hacia el encéfalo hasta llegar a las regiones de la corteza cerebral correspondiente. Por ejemplo, las sensaciones de tacto y dolor llegan hasta la corteza parietal después de hacer un relevo en el tálamo. Las órdenes motoras se originan en la corteza frontal en las células piramidales. Algunas de estas neuronas poseen axones suficientemente largos que les permite llegar hasta la sustancia gris de la médula espinal para estimular directa o indirectamente la actividad de las motoneuronas (DeFelipe, 1992:563-607).
Estructura de una neurona
Una neurona se compone básicamente de un cuerpo celular (también llamado ‘soma’) a partir del cual se forman dos tipos de procesos celulares alargados: dendritas y axones. Las dendritas son más gruesas y cortas mientras que el axón es muy delgado y en algunas neuronas puede ser lo suficientemente largo hasta alcanzar varios centímetros. Cada uno de estos axones termina en una dilatación microscópica (botón sináptico) que contiene un neurotransmisor (Figura 3). La mayoría de contactos sinápticos ocurren sobre las dendritas debido a que estas representan más del 90% de la superficie neuronal. Una neurona recibe un sinnúmero de contactos que provienen de muchas otras neuronas. A su vez el axón de cada neurona se ramifica para conectarse con varias neuronas simultáneamente. Cada rama termina en un botón sináptico desde donde se libera el neurotransmisor correspondiente que puede tener un efecto excitador, inhibidor o neuromodulador sobre la neurona postsináptica (la que recibe el impulso).

Figura 3. Esquema de las partes de una neurona. Dentro del axón se encuentran los microtúbulos rodeados por moléculas de dineína. Los microtúbulos son como ‘rieles’ indispensables para llevar a cabo cualquiera de los dos tipos de transporte axonal. La dineína aporta la energía para el transporte axonal.
Transporte axonal
Dentro del cuerpo neuronal la célula sintetiza diferentes elementos y sustancias; parte de ellos son conducidos luego hasta las dendritas y axones. El transporte de sustancias desde el soma hasta el botón sináptico se conoce con el término de transporte axonal anterógrado (Figura 3). Los primeros indicios de este fenómeno fueron detectados hacia mediados de la década de los años 40 cuando Paul Weiss y sus colaboradores observaron la formación de dilataciones en el sitio donde fueron ligados nervios de las extremidades posteriores. Luego de soltar la ligadura los nervios recobraron su diámetro normal. Concluyeron que las protuberancias formadas en los nervios ligados eran el resultado de la acumulación de material celular lo cual hacía suponer la existencia de un mecanismo de transporte activo de sustancias a lo largo de los axones y no una simple difusión pasiva (Weiss y Hiscoe, 1948:315-395). Investigaciones posteriores revelaron los detalles celulares y moleculares de este mecanismo de transporte.
Ahora bien, no resulta extraño aceptar que los componentes producidos en el cuerpo neuronal ‘viajen’ hasta lugares distantes de la neurona como son los botones sinápticos. Por el contrario, es más difícil aceptar que una molécula o partícula se desplace en dirección contraria, es decir, desde la terminal sináptica hasta el soma neuronal. Esto fue lo que unos investigadores del Instituto Karolinska (Estocolmo) reportaron en la década de los años 70 (Kristensson y Olsson, 1971:363-385). A este evento se le conoce como transporte axonal retrógrado (Figura 3), una especie de transporte en ‘contravía’ que requiere gran gasto de energía por parte de las neuronas para conducir sustancias hasta el citoplasma neuronal que luego son utilizadas en diferentes procesos metabólicos.
Transporte axonal retrógrado del virus de la rabia
En 1979 Henri Tsiang, entonces investigador del Instituto Pasteur en Paris, demostró experimentalmente que el virus de la rabia viaja a lo largo de los axones por transporte axonal retrógrado. Utilizó colchicina aplicada al nervio ciático de ratas previamente inoculadas con el virus en las plantas de sus patas traseras y logró detener el avance de la infección. La colchicina destruye los microtúbulos presentes en los axones lo que impide el transporte axonal en cualquier sentido. El uso de colchina marcada con tritio permitió establecer que se trataba de transporte axonal retrógrado (Tsiang, 1979:286-296). Estudios posteriores, entre ellos los de Gabrielle Ugolini (CNRS, Paris) quizás la investigadora que mejor ha estudiado el tema, ratifican que el mecanismo de transporte axonal retrógrado es el más común y más eficiente para que el virus de la rabia ingrese desde los tejidos periféricos donde es inoculado a través de la saliva de un animal infectado. De acuerdo con Ugolini, el virus ingresa a través de las terminaciones sinápticas que hacen contacto con las fibras musculares. Estas sinapsis corresponden a los axones que se originan en las neuronas motoras localizadas en el asta ventral de la médula espinal. Las motoneuronas espinales a su vez reciben contactos de axones que proceden de neuronas localizadas en niveles superiores del sistema nervioso, especialmente del tallo cerebral y la corteza cerebral frontal. Lo que se ha demostrado hasta ahora es que el virus sólo pasa de una neurona a otra por los sitios de contacto sináptico, desde la superficie del cuerpo celular o las dendritas de la neurona inicialmente infectada hasta el botón sináptico de la neurona aferente y desde allí se dirige hasta el cuerpo celular de la neurona que lo origina (Ugolini, 2011:165-202). Por lo tanto, una partícula viral que ingresa a través de una placa neuromuscular (el sitio de contacto entre un axón motor y la fibra muscular) puede llegar hasta la corteza cerebral haciendo como máximo 3 o 4 relevos, es decir, utilizando la ruta trazada por 3 o 4 neuronas.
Nosotros estamos estudiando la ruta neuroanatómica de ascenso del virus de la rabia inoculado en las extremidades posteriores de ratones gracias a la financiación de Colciencias y el Instituto Nacional de Salud y con el apoyo de la ACAC (Convenio INS-Colciencias-ACAC, 639 de 2014. Proyecto: “Estudio neuroanatómico de las dinámicas de infección celular en médula espinal y corteza cerebral en ratones inoculados con el virus de la rabia“). Hemos demostrado que el virus inoculado en los músculos isquiotibiales (muslo) de ratones se detecta mediante inmunohistoquímica inicialmente en motoneuronas de la médula espinal lumbar a las 30 horas (Figura 4), luego en neuronas motoras de la médula toráxica a las 40 horas y en la médula cervical a las 48 horas; desde allí el virus ascendió hasta las neuronas piramidales de la capa V de la corteza frontal motora en donde fue detectado inicialmente a las 64 horas luego de inoculado (Figura 5). Esta ruta señalada por el virus no deja duda de su preferencia por diseminarse, desde la periferia hasta la médula espinal, y luego al encéfalo, por transporte axonal retrógrado y a través de la vía motora (Torres-Fernández y otros, 2015:113-114).
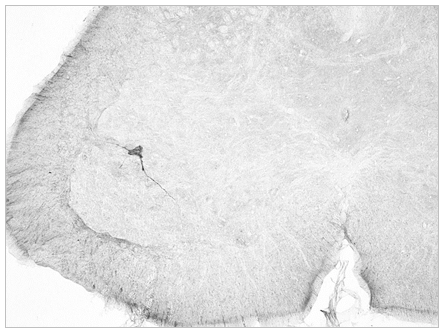
Figura 4. Imagen de una motoneurona (flecha) localizada en la médula lumbar 30 horas después de haberse inoculado virus de la rabia en el muslo de un ratón. Los antígenos llenan el citoplasma o cuerpo neuronal y parte de sus dendritass; su distribución se puede hacer visible mediante una reacción inmunohistoquímica.

Figura 5. Fotografía de un corte coronal (frontal) de encéfalo de ratón que revela la presencia de antígenos virales en neuronas piramidales de la capa V de la corteza cerebral, 72 horas después de la inoculación de virus de la rabia en sus extremidades posteriores.
Implicaciones del mecanismo de transporte del virus de la rabia
El hecho de que el virus de la rabia ingrese y se disperse exclusivamente a través de las vías trazadas por las conexiones neuronales lo protegen de la acción del sistema inmunológico. Por esta razón, una vez que el virus ha ingresado al interior de los axones de los nervios periféricos su dispersión hacia el resto del sistema nervioso es inatajable y el desenlace fatal inevitable. En neuronas en cultivo la velocidad del transporte axonal retrógrado del virus de la rabia se ha calculado en un rango de 50 a 100 mm por día (Tsiang y otros, 1991:1191-1194). In vivo esta velocidad parece ser inferior; así, por ejemplo, nosotros después de inocular ratones con virus de la rabia CVS (un tipo de virus de laboratorio) en los músculos isquiotibiales (localizados en la cara interior de los muslos), hemos detectado la llegada de los primeros antígenos de la rabia en las motoneuronas correspondientes dentro de la médula lumbar hacia las 30 horas postinoculación (Figura 4). La longitud de estos nervios motores entre la médula espinal y los músculos isquiotibiales en ratón es de unos 2,5 cm (25 mm), por lo tanto, se podría calcular la velocidad de desplazamiento del virus en 1,2 mm por hora (29 mm por día). El virus experimental CVS está altamente adaptado al modelo de ratón y por esta razón infecta al sistema nervioso del roedor casi inmediatamente después de ser inoculado por vía intramuscular (Ugolini, 2011:165-202). No obstante, en condiciones naturales el desarrollo de la enfermedad puede tardar algún tiempo luego de ser inoculado el virus en el tejido muscular de un receptor. Por ejemplo, en seres humanos, la aparición de los primeros síntomas puede tardar algunas semanas o meses (Tsiang, 1993: 375-412). Esto se debe a que el virus puede permanecer ese tiempo ‘latente’ o replicándose en el tejido inicialmente infectado (músculo o piel) sin ingresar al sistema nervioso. Este es el tiempo durante el cual el tratamiento con suero antirrábico y la vacuna lo pueden neutralizar.
Uso del virus de la rabia como trazador neuronal
Los trazadores neuronales son sustancias que se pueden inyectar en cualquier lugar del sistema nervioso para luego ser conducidas a otras áreas ya sea por transporte axonal retrógrado o anterógrado. Los mejores trazadores son los que sólo se transportan en una dirección (Lanciego y otros, 1999:24-28). Uno de los primeros compuestos hallados con esta propiedad fue la peroxidasa de rábano, un trazador neuronal retrógrado. Si este trazador es inyectado en nervios periféricos de los músculos de las extremidades posteriores de ratones es luego detectado mediante una reacción histoquímica en las neuronas piramidales de la capa cinco de la corteza cerebral. Esto mismo ocurre si se inyecta virus de la rabia en los músculos isquiotibiales de un ratón y luego, a partir de las 64 horas, se sacrifica el animal, se hacen cortes frontales (coronales) de la corteza cerebral y se revela la presencia de los antígenos virales mediante inmunohistoquímica (Figura 5). Esta es una forma didáctica de mostrar el transporte axonal retrógrado del virus rábico y la función motora de las neuronas piramidales de la corteza frontal. Diferentes laboratorios de neuroanatomía en el mundo están utilizando cepas no infecciosas o recombinantes del virus de la rabia como trazadores neuronales.
El virus de la rabia tiene ventajas sobre los trazadores químicos convencionales. Una de ellas es que parte del contenido de los trazadores puede difundirse fuera del territorio delimitado por las neuronas; esto a veces crea algo de confusión sobre el recorrido exacto de una vía neuroanatómica. Por el contrario, el virus de la rabia pasa de una neurona a otra únicamente a través de la unión sináptica, desde la neurona postsináptica hacia la neurona presináptica. Ugolini sostiene que no hay difusión o transporte del virus de una neurona a otra a través de otros contactos célula-célula (ya sean entre neuronas o con células gliales). El herpes, otro virus neurotrópico de transporte retrógrado no es tan específico, puede diseminarse de una neurona a otra por cualquier contacto neurona a neurona y fácilmente infecta células gliales, mientras que la infección glial con rabia es escasa. Otra ventaja del virus de la rabia como trazador es que se reproduce y va pasando de neurona a neurona por tiempo indefinido mientras que el trazador químico se va agotando, por lo tanto, tienen un alcance limitado a dos o máximo tres relevos neuronales Por estas razones, se considera al virus de la rabia como el mejor trazador neuronal que existe actualmente (Ugolini, 2011:165-202).
Es importante profundizar en el conocimiento que se tiene sobre los mecanismos de transporte del virus de la rabia. Quizás ahí podría estar el secreto para hallar compuestos viricidas que puedan ‘perseguir’ al virus y bloquear su avance en ‘contravía’ dentro de las vías neuroanatómicas. Por el conocimiento acumulado que se tiene sobre la patogénesis de la rabia se sabe que el virus primero asegura su diseminación a través de todo el sistema nervioso antes de que el hospedero manifieste signos de enfermedad. Por ejemplo, el electroencefalograma sólo refleja cambios en fase avanzada de la enfermedad. Y a nivel celular se ha demostrado que las neuronas en cultivo infectadas con el virus tardan un tiempo en mostrar alteraciones fisiológicas (Tsiang, 1993: 375-412). Nosotros hemos encontrado en el modelo animal que la expresión de algunas proteínas neuronales sólo se afecta en fases avanzadas de la enfermedad (Lamprea y Torres-Fernández, 2008: 7-13). Es como si el virus nos estuviera retando a neutralizarlo dentro de las neuronas antes de iniciar el daño fisiopatológico irreversible.
Referencias
Dean, D.J., Evans, W.M., McClure, R.C. (1963), “Pathogenesis of rabies”, en Bulletin of the World Health Organization, Vol. 29, pp. 803-811.
DeFelipe, J. (1992), “The pyramidal neuron of the cerebral cortex: morphological and chemical characteristics of the synaptic imputs”, en Progress in Neurobiology, Vol. 39, pp. 563-607.
Jackson, A.C. (2013), “History of rabies research”, en Jackson, A.C., (Ed.) Rabies: Scientific basis of the disease and its management, pp. 1-15, San Diego, Academic Press.
Kristensson, K., Olsson, Y. (1971), “Retrograde axonal transport of protein”, en Brain Research, Vol. 29, pp. 363-365.
Lamprea, N., Torres-Fernández, O. (2008), “Evaluación inmunohistoquímica de la expresión de calbindina en el cerebro de ratones en diferentes tiempos después de la inoculación con el virus de la rabia”, en Colombia Médica, Vol. 39 (Supl. 3), pp. 7-13.
Lanciego, J.L., Mengual, E., Erro, E., Giménez-Amaya, J.M. (1999), “Trazadores neuronales y su uso combinado”, en Revista Médica de la Universidad de Navarra, Vol. 4, pp. 24-28.
Schindler, R. (1961), “Studies on the pathogenesis of rabies”, en Bulletin of the World Health Organization, Vol. 25, pp. 119-126.
Torres-Fernández, O., Santamaría, G., Monroy-Gómez, J. (2015), “Dinámica neuroanatómica de infección celular en la ruta de dispersión del virus de la rabia en ratones inoculados por vía intramuscular”, en Biomédica, Vol. 35 (Supl. 3), pp. 113-114.
Tsiang, H. (1979), “Evidence for an intraaxonal transport of fixed and street rabies virus”, en Journal of Neuropathology and Experimental Neurology, Vol. 38, pp. 286-296.
Tsiang, H. (1993), “Pathophysiology of rabies virus infection of the nervous system”, en Advances in Virus Research, Vol. 42, pp. 375-412.
Tsiang, H., Ceccaldi, P. E., Lycke, E. (1991), “Rabies virus infection and transport in human sensory dorsal root ganglia neurons”, en Journal of General Virology, Vol. 72, pp. 1191–1194.
Ugolini, G. (2011), “Rabies virus as a transneural tracer of neural connections”, en Advances in Virus Research, Vol. 79, pp. 165-202.
Weiss, P., Hiscoe, H.B. (1948), “Experiments of the mechanism of nerve growth”, en Journal of Experimental Zoology, Vol. 107, pp. 315-395.






 NOTICIAS
NOTICIAS

